Evolution des tierischen Körperbaus
Kanonische Wnt- und Bmp/Chordin-Signalübertragung bei der Achsenbildung und -regeneration
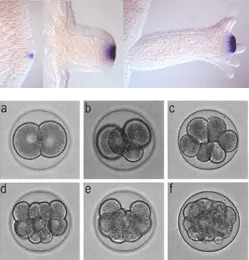
Nesseltiere verfügen über einen hoch konservierten kanonischen Wnt-Weg. Beim Hydrozoenpolypen Hydra stellt dieser Weg ein Schlüsselelement im Kopforganisator dar, der die Positionsinformationen entlang der Hauptachse zwischen Mund und Mundhöhle und während der Regeneration steuert. Im Polypen Nematostella werden Wnts während der Gastrulation im blastoporalen Organizer aktiviert, der sich später zum Kopf der Polypen entwickelt. Bei beiden Arten können wir ektopische Kopfstrukturen induzieren oder die Kopfbildung mit Hilfe von niedermolekularen Inhibitoren gegen GSK3 bzw. beta-Catenin hemmen. Unsere Studien zielen auf ein umfassenderes Verständnis des molekularen Netzwerks ab, das in diesen Organismen wirkt. Unsere früheren Ergebnisse deuten auch darauf hin, dass das Bmp/Chordin-Signalsystem bei der Musterung der oral-aboralen Achse von Nesseltieren eine Rolle spielt, wobei Chordin in den oralen Organisatoren aktiviert wird. Bei Bilateralen reguliert dieses Signalsystem die Bildung der dorsal-ventralen Achse. Um den Ursprung der Körperachsen der Bilateria aus eher radialsymmetrischen Vorfahren zu ergründen, haben wir begonnen, die Wnt- und Bmp/Chordin-Signalwege in den basalen bilateralen Plattwürmern Macrostomum lignano (Rhabditophora) und Isodiametra pulchra (Acoelomorpha) zu untersuchen.
Nicht-kanonische Wnt-Signale und Aktindynamik während der Gewebeevagination
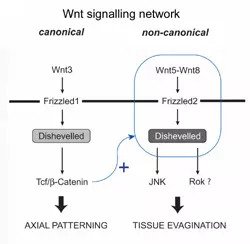
Basale Eumetazoen waren die ersten Tiere, die komplexe morphogenetische Gewebebewegungen erfanden, die zu Körperauswüchsen und Anhängseln führten. Um die molekulare Grundlage der Gewebeevagination bei Hydra, einem basalen Eumetazoen, zu verstehen, haben wir einen nicht-kanonischen Wnt-Signalweg charakterisiert, der Wnt5, Wnt8, Frizzled2, JNK, Rho-Kinase und Strabismus umfasst. Dieser Signalweg wirkt bei konvergenten, streckungsähnlichen Gewebebewegungen während der Knospen- und Tentakelbildung. Interessanterweise wird der nicht-kanonische Wnt-Signalweg in Hydra direkt durch den kanonischen Wnt-Weg aktiviert. Diese Koordination scheint wesentlich zu sein, um den korrekten Polypenkörperplan zu definieren, bei dem sich Tentakel und Knospen an den entsprechenden Positionen entlang der oral-aboralen Körperachse ausbilden. In Zukunft wollen wir die molekulare Interaktion zwischen dem kanonischen und dem nicht-kanonischen Wnt-Signalweg in Nesseltieren genauer analysieren. Darüber hinaus planen wir, die Aktin-bindenden Proteine zu charakterisieren, die an der Zellpolarisierung während der Knospen- und Tentakelbildung beteiligt sind, um einen besseren Einblick in die Mechanismen der Gewebeevagination zu erhalten.
Zelladhäsion auf Cadherin-Catenin-Basis und die Entstehung von echten Epithelien
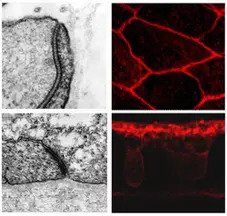
Die Entwicklung echter Epithelien war eine wichtige Erfindung in der Phylogenie der Eumetazoen (Hyman: "animals of tissue grade"). Epithelzellen weisen eine apikal-basale Polarität, apikale Zell-Zell-Verbindungen und Zellkontaktstellen mit einer basalen extrazellulären Matrix auf. Hydra ist ein hervorragendes experimentelles System, um die Entwicklung von Zelladhäsion und Epithelgewebe zu untersuchen, da es Zugang zu einer vollständigen Selbstorganisation in Reaggregationsversuchen bietet, die von einzelnen Zellsuspensionen ausgehen. In der jüngeren Vergangenheit haben wir die wichtigsten Mitglieder eines Cadherin-Catenin-Zelladhäsionskomplexes in Hydra charakterisiert. Wir haben auch desmosomale Verbindungen in Epithelzellen in Hydra identifiziert. All diese Daten unterstützen die Ansicht, dass Nesseltiere über einen funktionellen klassischen Cadherin-Catenin-Adhäsionskomplex verfügen, der an der Herstellung von Zell-Zell-Kontaktstellen beteiligt ist. Derzeit versuchen wir, die genaue intrazelluläre Lokalisierung dieser Proteine mit Hilfe von Immunogold-Färbungen in der Transmissionselektronenmikroskopie zu bestimmen. Langfristig wollen wir verstehen, wie Zelladhäsion und induktive Signalübertragung bei der Selbstorganisation zusammenwirken.
Mitglieder dieser Arbeitsgruppe
- Bert Hobmayer (head)